|
|
|
 |
Pioneering Work on Mistaken Point Fauna |
 |
 |
 |
 |
Fossils of the Precambrian Conception Group in the Biscay Bay-Cape Race area, Southeastern Newfoundland, are found as hundreds of imprints on the top surfaces of the graded beds of turbidite origin. the imprints are grouped into four categories:
The fossils are believed to be the imprints of soft-bodied Metazoans (Coelenterates) represented by Polyps as well as Medusae. The Polyps may have affinity either to Pennatulid Corals or they may represent new forms of colonial Hydrozoans. Although there is no evidence for ascertaining the age of the fossils, the fossil-bearing rocks are regarded as Precambrian in age based on lithologic correlation.[TOP]
Fossils in the Conception Group were discovered by the author (Misra, 1969) in the Biscay Bay-Cape Race area (Fig. 1) during the summer of 1967. A preliminary account of the Conception Group and a general discussion on its age has already been given (Anderson and Misra, 1968). Although only one category of impressions was reported in the 1968 communication, three other new categories have been found at the same locality. Several sub-categories have also been distinguished.
Prior to the discovery of Precambrian fossils in the Biscay Bay-Cape Race area, there was no record of life in the Precambrian rocks of Newfoundland, except for the doubtful fossil Aspidella terranovica Billings from the St. John's Formation. Indeed, throughout the whole North American continent there are only a few Precambrian fossil occurrences (Bassler, 1941;)Van Gundy, 1951; Frarey and McLaren, 1963). Most are doubtful except the occurrence of an apparent jelly fish imprint from the Grand Canyon, Arizona The fossil was discovered by C.E.Van Gundy in 1935 and was tentatively classified as Brooksella canyonensis (Basler, 1941). The most important Precambrian fossil locality thus far reported is in the Ediacara hills in southern Australia. It was discovered by an Australian geologist, R.C. Sprigg, during field studies in 1947 (Glaessner, 1961). The fauna of the Ediacara hills includes jelly fish, soft corals related to Pennatulids, segmented worms, and other animals (Glaessner, 1961). Most of these organisms belong to the Phylea Coelenterata and Annelida. Precambrian fossils are known also form South Africa and England (Glaessner, 1959).
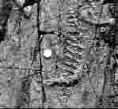
The fossils of the Conception Group occur as hundreds of imprints on ripple-marked surfaces of graded beds consisting of graywackes at the bottom, through siltstone and argillites at the top. The fossils have been observed at several horizons within a thickness of about 175 feet.The fossil-bearing planes are generally overlain by about 5 inch thick volcanic (?) tuff (Pls. 2 and 3, dark areas. It was difficult to sample the material, and studies on the covering layer are not conclusive. The fossil-bearing rocks are exposed only on the coast; lateral extension of the beds inland is not known. Investigation at the projected same stratigraphic level at Cape Cove near Cape Race (Fig. 1) did not give fruitful results. It is probable that population of organisms was most dense near Mistaken Point and although lateral and vertical extensions of the fossil-bearing beds and change in the fossil characters with time are not known, the concentration of animal impressions in a limited area suggests that these soft-bodied Metazoans constituted a flourishing fauna during the time of sedimentation of the Conception Group. [TOP]
Click here to See Plate Section for all plates.
The number of main fossil categories is small compared with the Ediacara fauna of southern Australia but the variations within each category are many. The detailed picture of these variations is as yet not clearly understood. However, it is believed that the fossils include Polyps as well as Medusae of Coelenterates, and have been grouped into four categories described later in this paper. Each fossil category is represented by numerous individuals, usually in random orientation. The fossils are found on exposed surfaces and those within the reach of sea waves have become worn. the surfaces which are either protected by the covering rock or are attacked only occasionally by the sea waves contain well-defined fossils with distinct and sharp details of their body structure
The argillites that contain the fossils are fractured and jointed (Pls.2,3, and 4 ) making it difficult to sample the fossils, but four specimens were collected in the field and studied in thin section The fossil-bearing argillites show no organic remains under the ordinary microscope. Studies under electron microscope or chemical studies under electron microscope or chemical studies might yield better results. In the absence of actual fossil specimens the present work is based on the photographs of the fossils and their casts exhibit raised branches in the case of spindle-shaped organisms (Pls. 3 and 8C) but the same parts look depressed if viewed after rotating the photograph by 180. Similar effect is seen also in the other categories, especially in the case of lobes in the round lobate organisms. (Pl. 4A). This three-dimensional effect is probably due to composite molding of parts of dorsal and ventral surface of the animals. thus, it is apparent that the organisms had three-dimensional bodies and were not flat-bodied organisms.
Since the fossils are found in a turbidite sequence of graded graywackes, siltstones, and argillites containing load casts, flute casts, and prod marks, the sessile forms of the fauna must have lived in a moderately deep environment. They were probably living on the bottom of the sea during the intervening quiescent period between two successive turbidity currents and lived until they were buried by sediments brought by suffocating turbidity currents and lived until they were buried by sediments brought by suffocating turbidity currents or were killed by mild volcanism (?) . However, if they represent floating organisms, they must have come to rest on the muddy bottom and made either imprints on the substrate or were entombed bodily. In either case, the surface of mud provided the ideal conditions for preserving imprints, and the fossil impressions, regardless of their shape and size, are clear and well-defined. The strata underlying and overlying the fossil-bearingbeds are unfossiliferous, suggesting the animals flourished and died in a short interval of time. All the them were softbodied organisms because no part of their body is found preserved. Also, the fossils it seems, are preserved in situ because the impressions of soft, delicate bodies after transport could not be expected to be found in entirety. [TOP]
I am grateful to Professor M.F. Glaessner of Adelaide University, South Australia for his help and encouragement. The fossils were found during a geological study of the Biscay Bay-Cape Race area for the Master of Science degree at Memorial University of Newfoundland, and I am indebted to Professor W.D. Brueckner for suggesting this thesis area. The study was partly financed by Professor Brueckner's national Research Council grant A-884 and partly by his Geological Survey of Canada grant. Research facilities provided at Memorial University of Newfoundland and helpful discussions with faculty members are also gratefully acknowledged.[TOP]
Most geological studies in the Avalon Peninsula of Newfoundland have been confined to the Torbay (Rose, 1952) and Whitbourne (McCartney, 1967) map areas. The area south of lat 470 has not been mapped . The Biscay Bay-Cape Race area (Fig. 1), which was mapped by the author during the summer of 1967, lies on the extreme southeastern part of the unmapped peninsula. The map area is underlain by a thick sequence of sedimentary rocks which can be separated by a gradational boundary into two main divisions: a lower division consisting of banded cherts, argillites, graywackes and siltstones corresponding to the Conception Group (Rose, 1952); and an upper division consisting of shales with sandstone laminae corresponding to the St.John's Formation of the Cabot Group (Rsse, 1952). The rocks of the lower division are mainly green and purple and those of the upper division are various shades of gray. The author has checked the correlation of these division with the Conception Group and the St. John's Formation by reconnaissance study in the intervening unmapped area from Portugal Cove south to Ferryland, and found that the lithologic correlation is clear and definite.
The rocks of the Conception Group in the Torbay map area (Rose, 1952) were found to overlie the Precambrian Harbour Main Group unconformably, and to underlie the St. John's Formation of the Cabot Group disconformably. In the Harbour Grace (Hutchinson, 1953) and Whitbourne (McCartney, 1967) map areas of the Avalon Peninsula, the Conception. Group is uncomfortably underlain by the Harbour Main Group and Holyrood Granite, and conformably overlain by the Carbonear Formation of the Hodgewater /Group. The Hodgewater Group has a thickness of about 10,000 feet and, except for the Random Formation, forms the top of the Precambrian sequence in the Harbour Grace area (Hutchinson, 1953). Thus,the Conception Group is underlain as well as overlain by the rocks that are believed to be Precambrian in age.
Non-mixing of early lower Cambrian diagnastic fauna in the "Atlantic" and "Pacific" faunal zones seems to present unique palaeontologic complexity. The Callavia and Protolenus faunal zones of eastern Newfoundland cannot be precisely correlated with Olenellus beds of the "Pacific" realm, so widespread elsewhere in North America (McCartney and other , 1966). The base of the Cambrian system is drawn by Hutchinson (1962) at the first disconformity in the stratigraphic sequence below the rocks of the Callavia zone, and the mean isochron age for base of the Cambrian in the Atlantic Provinces of Canada is 500 � 40 m.y. (Fairbairn and other, 1966). This figure when compared with the standard time scale (Cowie, 1964; Glaesquestion can, therefore, be raised if we are dealing with two different time scales in the early Cambrian (McCartney and other, 1966), and if the term. Precambrian signifies pre-Palaeozoic in the sense of organic evolution and not pre-Cambrian in the chronologic sense (Cloud and Nelson, 1966).
The age of the Conception Group is not precisely known. The age determination was made by the Geological Survey of Canada (G.S.C. paper 66-17, 118 p.) on a small specimen of orthoclase from lava in the Conception Group. The analysis indicates a potassium-argon age of 457�82m.y. However, McCartney (1967) is of the opinion that the age should be near the older extremity of the inferred margin of error. Nevertheless, the age is questionable because the top of the Conception Group lies at least 14,000 feet below the Precambrian-Cambrian boundary in the Torbay map area (Rose, 1952) 9000 feet below in the Whitbourne map area (McCartney, 1967), and 10,000 feet in the Harbour Grace map area (Hutchinson, 1953).
The age of the Harbour Main Group, which directly underlies the conception Group. is 568�29 m.y. (Fairbairn and others, 1966). However, its contact relation to the Conception Group is uncertain. It is possible that the lower part of the Conception Group is partly synchronous with the youngest rocks of the Harbour Main Group. Also, the age of the Holyrood Granite, which intrudes the Harbour Main and the Conception Groups, is 574�11m.y. (McCartney and other, 1966). The author does not know why the age of the intrusive is more than that of the host rocks because both figures represent Rb/Sr age. The problem of the age of the Conception group may be resolved after the evolutionary history of the Metazoans is clearly understood. However, the age considerations of the underlying and intrusive rocks together with the stratigraphic position suggest a Precambrian age of the Conception Group, probably very late Precambrian in the chronologic sense. [TOP]
The fossils are preserved as impressions and can be grouped into the following four categories:
These are the commonest categories of the fossil impressions, which also have variations as discussed in the following pages. A category includes the organisms that exhibit similarity in their body structure, and may include more than one genera. A descriptive terminology is not proposed in this paper because of uncertainties involved in the identification of the organisms [TOP]
Plates 1B, 1C, 1E; 2A,2C; 3A, 3C; 4C; 5A, 5C; 6C; 7B, 7C, 7D; 8C,8D.
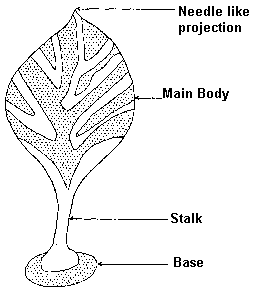 |
On the basis of visual reservation, these fossils are second in abundance next to the spindle-shaped organisms. A leaf-shaped organism consists of three parts: a main body having leaf-shaped structure (Pls. 3B and 6D) and needle-shaped projection (Pls. 1D,7A, and Fig. 2) ; an stalk (Pls. 4B and 6D); and a round base attached to the stalk (Pl. 4B). The fossils are oriented roughly in the direction of the turbidity currents with the main body always falling in the direction of flow (Pl. 2B). The animals in some cases were broken from the base and moved slightly in the direction of the currents, leaving the disc-shaped base behind (Pl. 1H); in other cases only the main body broke away and the base and the stalk are found together (Pl.1G). |
The main body of some animals was compressed and their width reduced in the fossil form (Pl. 5B)k, while others are found with their body intact (Pls. 1D, 6D, and 8A). The sizes of different parts of the organisms were measured for 10 specimens and these measurements suggest that the specimens represent an ontogenic series. Some organisms have a proportionately longer stalk (Pl. 4B) while others have a small stalk but a large body (Pl.1D) Some specimens are worn out and their body structure is obscured (Pl. 3D). Details of the fossil are not clear in the casts but the fossil are not clear in the casts but the positions of base, stalk, and the main body are evident (Pl. 8A).
Plates 1A,1F,2D,3E,4A,5D, and 6B.
Distribution and Form
The outline of these organisms is distinct and their form irregular. In smaller organisms of the sub-category, the peripheral lobes are attached to a lobe in the centre (Pl. 6B). However, in the larger specimens this relation is indistinct (Pl. 4A). Some of the larger organisms exhibit a net-like structure with a depression in the central part (Pl. 1F). Some other organisms included in the sub-category representing, presumably, more than one organism attached to each other.
The fossils are found only as impressions, suggesting they were soft-bodied Metazoans that constituted a flourishing fauna during the late Precambrian times in the Conception Sea. It is not yet clear if and to what extent this Precambrian fauna of Newfoundland is related to the Edicara fauna of southern Australia. It is probable that spindle-shaped, leaf-shaped, and dendrite-like organisms may be Pennatulid corals and may be compared with other Precambrian faunas of the world. However, it is also possible that the fossils represent an entirely new fauna heretofore unreported. nevertheless, some of the lobate organisms may have affinity to Brooksella canyonensis (Bassler, 1941) reported from Arizona. Based on branching of the spindle-shaped organisms, an alternative explanation is that the organisms are colonial floating Hydrozoans belonging to the division Calyptoblastia. The round lobate impressions may be floats as suggested by the base of leaf-shaped objects, or Medusae. The dendrite-like forms may be the same as spindles in a different preservation, and the leaf-shaped structures may be related[TOP]
Revised Manuscript Received April 21. 1969.
